This is the translation. The original web-page (oryginalna strona): http://www.tulane.edu/~wiser/protozoology/notes/INTRO.html
Mark F. Wiser
Co to są pierwotniaki?
Odszyfrowanie greckich korzeni skutkuje zdefiniowaniem pierwotniaków jako „pierwszych” (proto) „zwierząt” (zoa). Chociaż molekularne badania filogenetyczne wskazują, że pierwotniaki należą do najwcześniej rozgałęziających się eukariotów (patrz drzewo filogenetyczne), taka definicja nie dostarcza wielu informacji opisowych. Pierwotniaki nie są łatwe do zdefiniowania, ponieważ są różnorodne i często są ze sobą tylko daleko spokrewnione. Ze względu na ogromną różnorodność pierwotniaków jedyną wspólną cechą wszystkich pierwotniaków jest to, że są one jednokomórkowymi mikroorganizmami eukariotycznymi. Pierwotniaki posiadają typowe organelle eukariotyczne i na ogół wykazują typowe cechy innych komórek eukariotycznych. Na przykład jądro związane z błoną zawierające chromosomy znajduje się u wszystkich gatunków pierwotniaków. Jednak u wielu gatunków pierwotniaków niektóre organelle mogą być nieobecne lub mogą różnić się morfologicznie lub funkcjonalnie od tych występujących u innych eukariotów. Ponadto wiele pierwotniaków ma organelle, które są unikalne dla określonej grupy pierwotniaków.
Pytania, które można by zadać w odniesieniu do definicji pierwotniaków, obejmują: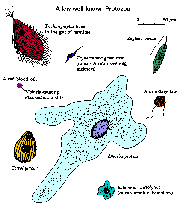
Pierwotniaki wykazują różnorodne morfologie (kliknij rysunek, aby powiększyć obraz i opis). Nie ma jednego kształtu ani morfologii, która obejmowałaby większość pierwotniaków. Kształty wahają się od amorficznych i ciągle zmieniających się form ameby do stosunkowo sztywnych form podyktowanych częściowo wysoce uporządkowanymi cytoszkieletami lub wydzielonymi ścianami lub muszlami. Kilka gatunków pierwotniaków wykazuje barwniki fotosyntetyczne lub inne, a zatem jest zabarwionych. Wiele gatunków pierwotniaków wykazuje złożone cykle życiowe z wieloma etapami. Czasami różne etapy cyklu życia są tak różne, że mylono je z zupełnie różnymi gatunkami.
Rozmiary pierwotniaków
| Organizm | Rozmiar |
|---|---|
| Merozoity Plasmodium | 2-5 μm |
| Pantofelek | 200-500 μm |
| Spirostomum ambiguum | 3 mm |
| Nummulity (skamieniałe) | 7-13 cm |
Pierwotniaki – z wyjątkiem kilku form kolonialnych – są organizmami jednokomórkowymi lub jednokomórkowymi; chociaż niektórzy twierdzą, że są one w rzeczywistości „bezkomórkowe”. Tak więc zdecydowana większość pierwotniaków jest mikroskopijna. Jednak wykazują niewiarygodnie szeroką gamę rozmiarów. Rozmiary ocalałych gatunków wahają się od <1 µm (10-6 metrów) do kilku mm. Zidentyfikowano skamieniałą forminiferida o wielkości kilku cm. (Wymarłe pierwotniaki można wykryć dzięki wydzielonej skorupce węglanu wapnia.) Większość omawianych w tym kursie organizmów będzie miała wielkość 3-50 µm. Ten niewielki rozmiar wymusza użycie mikroskopu do wykrywania pierwotniaków. Do szczegółowych badań morfologicznych potrzebny jest mikroskop elektronowy.
Pierwotniaki występują w wilgotnym środowisku praktycznie wszędzie. Jako grupa pierwotniaki są niezwykle elastyczne. Poszczególne gatunki mają jednak na ogół specyficzne nisze. Podobnie jak wszystkie inne organizmy, pierwotniaki muszą być w stanie pobierać i metabolizować składniki odżywcze ze swojego środowiska (tj. heterotroficzne). Wiele pierwotniaków po prostu absorbuje substancje rozpuszczone (tj. osmotrofię) ze swoich pożywek, podczas gdy niektóre są zmiataczami, które połykają stały materiał (tj. fagotrofia). Drapieżne pierwotniaki albo aktywnie polują na inne organizmy (zwykle bakterie lub inne pierwotniaki), albo biernie je zasadzają. Niektóre pierwotniaki są fotosyntetyczne i mogą wychwytywać energię słońca i przekształcać ją w użyteczną energię chemiczną (tj. autotroficzną lub fototroficzną). Wiele pierwotniaków nie ogranicza się do jednego mechanizmu żywienia i może wykorzystywać kombinacje powyższych (tj. miksotroficzne).
Pierwotniaki można również postrzegać jako żyjące na wolności lub symbiotyczne. Generalnie organizmy wolno żyjące występują w glebie lub w środowisku wodnym, podczas gdy symbionty żyją w bliskim związku z innym organizmem. Symbioza oznacza fizjologiczną zależność jednego organizmu od innego organizmu, a nie tylko ścisły związek fizyczny między dwoma organizmami. Generalnie ta zależność ma formę odżywiania. Można wyróżnić różne formy symbiozy, które odzwierciedlają naturę związku między dwoma organizmami (ramka).
Skojarzenia symbiotyczne
| Komensalizm | Oznacza interakcję korzystną dla jednego organizmu, ale nie mającą wpływu na inny organizm. Na przykład wiele pierwotniaków żyje w przewodzie pokarmowym innego organizmu, nie szkodząc mu. Te komensały są często padlinożercami lub drapieżnikami, które wykorzystują obfitość składników odżywczych lub fauny bakteryjnej dostarczanej przez organizm żywiciela. |
| Wzajemność | Oznacza specjalną formę komensalizmu, w której oba organizmy czerpią pewne korzyści i stają się od siebie zależne. Klasycznym przykładem mutualizmu jest pierwotniak Trichonympha znaleziony w jelitach termitów. Trichonympha przy pomocy symbiotycznej bakterii trawi cząsteczki drewna (tj. Celulozę) wchłonięte przez termit. |
| Pasożytnictwo | Oznacza związek, w którym jeden organizm (pasożyt) odnosi korzyści kosztem innego organizmu (żywiciela). Na ogół ten wydatek żywiciela oznacza, że pasożyt przyjmuje makrocząsteczki od żywiciela i wypuszcza inne do żywiciela. W niektórych przypadkach pasożytnictwo będzie jawnie szkodliwe dla żywiciela i określane jako chorobotwórcze. Przedmiotem tego kursu są przede wszystkim te chorobotwórcze pierwotniaki. |
Ruchliwość i cytoszkielet
| Tryby ruchliwości | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
Najwcześniejsze obserwacje pierwotniaków wskazywały na ich ruchliwość. Ta ruchliwość skutkowała ich klasyfikacją jako „zwierzęta”, które odróżniono od nieruchliwych „roślin”. Jednak ruchliwość nie jest uniwersalną cechą pierwotniaków, a różne pierwotniaki wykorzystują różne mechanizmy ruchu (tabela). W rzeczywistości pierwotniaki zostały początkowo sklasyfikowane w oparciu o ich mechanizm ruchliwości (patrz Taksonomia).
Rzęski i wici to struktury subkomórkowe, które napędzają pierwotniaki przez płynny ośrodek. Wici to długie, przypominające bicze struktury, które wprawiają organizm w ruch w wyniku falowego uderzenia, które rozchodzi się na całej ich długości. Wiciowce mają zazwyczaj jedną lub kilka wici na organizm. Natomiast pierwotniaki orzęskowe są zwykle pokryte rzędami licznych rzęsek. Dudnienia tych rzęsek są skoordynowane i działają jak wiosła, napędzając organizm. Rzęski i wici mogą również pomagać w pozyskiwaniu pożywienia, rozmnażaniu i innych funkcjach. Rzęski i wici składają się z tych samych składników białkowych i są w rzeczywistości strukturami równoważnymi. Obie są wiązanymi z błoną włóknistymi wypustkami z komórki. Włókno, znane jako aksonem, składa się z szeregu równoległych mikrotubul, zazwyczaj wykazujących układ „9 + 2”. Ruch powstaje, gdy mikrotubule przesuwają się obok siebie. Siła, która pośredniczy w tym ruchu ślizgowym, jest generowana przez białko zwane dyneiną. Dyneiny to „białka motoryczne”, które przekształcają energię chemiczną uwolnioną podczas hydrolizy ATP w energię mechaniczną. Mikrotubule to elementy cytoszkieletu, które również odgrywają ważną rolę w kształtowaniu komórek i są głównym składnikiem wrzeciona mitotycznego.
W przeciwieństwie do pływania wykazywanego przez wiciowce i orzęski, ameba to pierwotniaki, które pełzają po twardym podłożu w sposób znany jako „ruch ameboidalny”. Ameba wystaje z ciała komórki pseudopodium lub fałszywą stopę. Pseudopodium następnie przyczepia się do podłoża, a następnie ciągnie resztę ciała komórki do przodu. Siła zaangażowana w ten ruch jest generowana przez inny układ cytoszkieletu, który składa się z aktyny i miozyny. Aktyna tworzy długie włókna, znane również jako mikrofilamenty, a miozyna jest białkiem motorycznym, które porusza się wzdłuż mikrowłókien w sposób zależny od ATP. Skurcz mięśni jest kolejnym przykładem generowania siły przez elementy cytoszkieletu aktynowo-miozynowego. W sensie mechanistycznym fagocytoza jest formą ruchu ameboidalnego obejmującego również mikrofilamenty. W tym przypadku pseudopodia są rozszerzone, aby otoczyć połkniętą cząstkę. Fuzja pseudopodiów z ciałem komórki powoduje internalizację cząstki w wakuoli.
Apicomplexa również czołgają się po podłożu, ale za pomocą innego mechanizmu niż ameba. Mechanizm tej tak zwanej „ruchliwości ślizgowej” dopiero zaczyna być rozumiany i prawdopodobnie obejmuje zarówno układy cytoszkieletowe oparte na mikrowłókien, jak i mikrotubulach. Apicomplexa wykazuje również formy wewnątrzkomórkowe, a inwazja komórki gospodarza również obejmuje tę ruchliwość ślizgową. (Zobacz także omówienie inwazji komórek gospodarza przez pasożyta malarii).
Ruchliwość komórkowa obejmuje generowanie siły przez elementy cytoszkieletu oparte na mikrotubulach lub elementy cytoszkieletu oparte na mikrotubulach. Dotyczy to pierwotniaków i innych eukariontów. Udział mikrotubul i mikrofilamentów zarówno w kształcie komórki, jak i ruchu komórek sprawia, że te struktury subkomórkowe są bardziej podobne do układu mięśniowo-szkieletowego.
Reprodukcja
Pierwotniaki, podobnie jak wszystkie inne organizmy, rozmnażają się. Najczęstszą formą
| Tryby reprodukcji | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|
|
rozmnażania pierwotniaków jest bezpłciowe rozszczepienie binarne. Innymi słowy, pojedynczy organizm podzieli się na dwa równe organizmy. Niewielka modyfikacja tego podwójnego rozszczepienia, zwanego pączkowaniem, występuje, gdy jedna z nowo powstałych komórek jest mniejsza od drugiej. Zwykle większa komórka nazywana jest matką, a mniejsza – córką. Niektóre pierwotniaki utworzą wewnątrzkomórkowy pączek i zasadniczo rodzą. Inną odmianą rozszczepienia binarnego jest rozszczepienie wielokrotne lub segmentacja. W tej sytuacji kilka rund replikacji jądra zachodzi bez cytokinezy. Ta wielojądrzasta komórka będzie następnie jednocześnie tworzyć wiele potomstwa.
Wiele pierwotniaków oprócz bezpłciowych form rozmnażania wykazuje także rozmnażanie płciowe. Ta rozmnażanie płciowe może obejmować produkcję i łączenie gamet w procesach podobnych do organizmów wyższych. Ciliophora przechodzi koniugację, w której przeciwne typy kojarzą się w pary i bezpośrednio wymieniają materiał genetyczny (tj. DNA). Czasami rozmnażanie płciowe jest obowiązkowym etapem cyklu życiowego, podczas gdy w innych przypadkach organizm może rozmnażać się bezpłciowo z okazjonalnymi cyklami rozmnażania płciowego.
Podsumowując, pierwotniaki to jednokomórkowe mikroorganizmy eukariotyczne. Jednak różnorodność pod względem morfologii, wielkości i stylu życia pierwotniaków utrudnia opracowanie dokładniejszej definicji. Ich długa historia ewolucyjna (patrz drzewo filogenetyczne) odpowiada za znaczną część tej różnorodności. Jednak pierwotniaki wykazują cechy wspólne dla wszystkich eukariontów. (Połączyć do serii prezentacji PowerPoint na temat biologii komórek eukariotycznych).
| Pochodzenie komórek eukariotycznych |
|---|
| Ewolucyjny skok od prokariontów do eukariontów jest znaczny i szacuje się, że istnieje 5000 genów znalezionych u eukariontów, których nie można znaleźć u prokariontów (1). Pierwsze organizmy eukariotyczne prawdopodobnie przypominały to, co teraz określilibyśmy jako pierwotniaki. Jednak prawdopodobnie obecnie nie istnieją żadne bezpośrednie potomki wczesnych eukariontów, a ścieżka od prokariontów do eukariontów jest prawdopodobnie zaśmiecona kilkoma brakującymi ogniwami. Dwa kluczowe wydarzenia w ewolucji eukariontów to prawdopodobnie rozwój wewnętrznego układu cytoszkieletowego i fagotrofia.Główną różnicą między eukariontami a prokariotami jest sztywna mureina lub peptydoglikan, ściana komórkowa bakterii. Chociaż wiele eukariontów wykazuje ściany komórkowe, są one zasadniczo inne niż bakterie i prawdopodobnie pochodzą wtórnie po uformowaniu się eukariontów. Na przykład ściany komórkowe komórek roślinnych składają się głównie z celulozy, a ściany komórkowe grzybów i cysty u wielu pierwotniaków składają się głównie z chityny. Sztywność i wytrzymałość zapewniane przez ściany komórkowe chronią komórki przed naprężeniami mechanicznymi i zapewniają stabilność osmotyczną. Zatem zbiegło się z utratą ściany komórkowej mureiny u wczesnych eukariontów, rozwój układów endocytowo-szkieletowych opartych na aktynie i tubulinie w celu kompensacji względnej kruchości nagiej błony plazmatycznej. Dodatkowo, wczesne eukarionty prawdopodobnie ewoluowały w środowisku bentosowym, w szczególności w gęstych morskich matach mikrobiologicznych. Organiczne substancje rozpuszczone wydzielane przez bakterie i wysokie stężenie soli w takim środowisku miałyby działanie osmoprotekcyjne. Gęste maty mikrobiologiczne zapewniałyby również środowisko, w którym ofiara była bardzo liczna, a fagocytoza mogłaby ewoluować bez wydajnej ruchliwości komórek.
Przodkowy eukariot prawdopodobnie przyczepił się do ofiary poprzez N-połączone glikoproteiny na powierzchni komórki, które wyewoluowały po utracie ściany komórkowej mureiny, a następnie strawił je zewnętrznie, wydzielając enzymy hydrolityczne. Aby uczynić ten proces bardziej wydajnym, prymitywne drapieżniki mogły stopniowo otaczać swoją ofiarę, aż ofiara została całkowicie pochłonięta przez błony żywiciela, a trawienie odbywało się wewnątrz fagosomów. Ten mechanizm endocytarny wymagałby również opracowania mechanizmu egzocytarnego, aby utrzymać powierzchnię błony komórkowej. Zatem ewolucja fagocytozy obejmowała nie tylko rozszerzenie funkcji cytoszkieletu aktynowego, ale także napędzała rozwój transportu błonowego i przedziałów subkomórkowych, w tym jądra. Ewolucja fagotrofii i endomembran nie tylko otworzyła nowy sposób odżywiania, ale także pozwoliła na endosymbiozę i zniewolenie mitochondriów. Wczesny eukariont prawdopodobnie pochłonął alfa-proteobakterię i utworzył symbiotyczny związek, który wyewoluował w asymilację metabolizmu tlenowego tego organizmu i rozwój mitochondrium. To zniewolenie mitochondrium było prawdopodobnie kolejnym kluczowym wydarzeniem w ewolucji eukariontów, ponieważ istniejące eukarionty pochodzą od przodków zawierających mitochondria. Istniejące beztlenowe eukarionty bez mitochondriów albo mają organelle pochodzące z mitochondriów, takie jak mitosomy i hydrogensomy, albo przynajmniej mają geny jądrowe kodujące pozostałe geny mitochondrialne. Przypuszczalnie amitochondrialny eukariotyczny przodek znajdował się w niekorzystnej sytuacji konkurencyjnej i nie przetrwał w perspektywie długoterminowej. Później cyjanbakterium zostało zniewolone przez przodka roślin i rozwinęło się w chloroplasty, podobnie jak poprzednicy tych roślin zostali pochłonięci i zniewoleni, tworząc plastydy występujące w wielu innych glonach i pierwotniakach. Cytoszkielet nadal ewoluował i rozszerzał swoje funkcje, aw szczególności zaczął odgrywać rolę w ruchliwości komórek. Przede wszystkim tubulina stała się kluczowym składnikiem wici, a później miozyna ewoluowała, aby umożliwić rozwój ruchów ameboidalnych i ruchliwości ślizgowej. Ta ruchliwość umożliwiła tym wczesnym eukariontom ekspansję do innych nisz (np. Planktonicznych, ziemskich), a zatem ten tlenowy, fagotroficzny i wiciowaty przodek eukariota mógł promieniować do różnorodnych pierwotniaków, a ostatecznie innych eukariotów, które obecnie istnieją.
|
Taksonomia
Taksonomia lub systematyka to nauka o nazywaniu i klasyfikowaniu organizmów. Oprócz przypisywania hierarchicznych klasyfikacji taksonomicznych, systematyka próbuje również umieszczać organizmy w grupach, odzwierciedlając relacje ewolucyjne lub filogeny. Jednak kryteria taksonomiczne są często arbitralne, a taksonomia zawsze się zmienia, aby odzwierciedlać nowe odkrycia i interpretacje. Ponadto cechy użytkowe, takie jak rodzaj choroby, zasięg żywiciela i rozmieszczenie geograficzne, są często wykorzystywane w systematyzacji mikroorganizmów chorobotwórczych. Jest to szczególnie prawdziwe w przypadku taksonomii pierwotniaków. Ponadto toczy się dyskusja na temat ogólnej filozofii klasyfikacji pierwotniaków, a związki między wieloma grupami pierwotniaków nie są znane (patrz D.J. Patterson w Am. Nat. 154, S96-124). Klasyfikację pierwotniaków dodatkowo komplikuje pierwotna definicja pierwotniaków jako jednokomórkowych heterotrofów. Obecnie uznaje się, że pierwotniaki (lub protisty) mogą wykorzystywać wiele strategii żywieniowych i nie można ich traktować jako po prostu roślinopodobne (autotrofy) lub zwierzęce (heterotrofy). Zatem terminu pierwotniaki nie można uznać za prawdziwą grupę taksonomiczną. Pomimo faktu, że samo słowo pierwotniak nie jest już właściwą nazwą taksonomiczną, nadal jest użytecznym i funkcjonalnym terminem.
Historycznie pierwotniaki podzielono na cztery główne grupy: ameba, wiciowce, orzęski i sporozoa. Cechami wyróżniającymi grupy była ruchliwość (tj. Ameboid, wici, rzęski). Zarodniki były heterogeniczną grupą, która produkowała zarodniki na jednym etapie swojego cyklu życiowego i wykazywała ruchliwość „ślizgową”. Jednak taki schemat klasyfikacji jest dość arbitralny i niekoniecznie odzwierciedla prawdziwe ewolucyjne relacje między organizmami. Jednym z problemów związanych ze stosowaniem ruchliwości jako kryterium taksonomicznym jest to, że wiele pierwotniaków wykorzystuje różne typy ruchliwości na różnych etapach swojego cyklu życiowego. Na przykład Naegleria występuje w formie ameby, gdy pożywienia jest pod dostatkiem i przekształca się w wiciokrzew, gdy brakuje pożywienia. Ogólnie rzecz biorąc, ameby są niejednorodną grupą i prawdopodobnie wszystkie pochodzą od wiciowców. Spośród tych czterech pierwotnych grup pierwotniaków tylko orzęski są nadal uważane za ważny takson.
Od lat sześćdziesiątych XX wieku mikroskop elektronowy był używany do identyfikacji cech ultrastrukturalnych, które mogą służyć jako kryteria grupowania pierwotniaków. W wielu przypadkach morfologia prowadzi do klasyfikacji, która dzieli organizmy na grupy monofiletyczne. Monofiletyczny oznacza, że wszystkie organizmy w tej grupie prawdopodobnie pochodzą od wspólnego przodka. Na przykład wiele pierwotniaków zwanych dawniej sporozoa posiada struktury subkomórkowe, zbiorczo zwane organellami wierzchołkowymi, a obecnie tworzy grupę monofiletyczną zwaną apicomplexa. Jednak struktury subkomórkowe i szlaki metaboliczne mogą zostać utracone w niektórych liniach, a umieszczanie tych potomków może być problamatyczne.
W latach osiemdziesiątych XX wieku i do dziś stosuje się techniki molekularne do taksonomii. Ewolucyjne historie i pokrewieństwa można wyprowadzić przez porównanie sekwencji DNA lub białek. Dane o sekwencjach molekularnych potwierdziły filogenezy oparte na innych kryteriach, rozstrzygnęły niektóre debaty i doprowadziły do kilku niespodzianek. Na przykład dane molekularne potwierdzają, że apikompleksy są monofiletyczne, a ponadto wskazują, że są one spokrewnione z orzęskami i bruzdnicami. Te trzy grupy są teraz połączone w większą grupę monofiletyczną o nazwie alveolata. Ten związek był już wcześniej podejrzewany, a nazwa nawiązuje do struktur morfologicznych znanych jako worki pęcherzykowe. Jednak użycie pojedynczych genów może być niewiarygodnym sposobem określenia relacji ewolucyjnych, szczególnie między odległymi organizmami, a dane molekularne należy interpretować z ostrożnością.
Od połowy lat osiemdziesiątych klasyfikacja protistów podlega ciągłym zmianom i jest dotknięta pewnymi filozoficznymi kontrowersjami. Z jednej strony istnieje argument przemawiający za zachowaniem elementów schematu Bütschliana opartego na ruchliwości ze względu na jego znajomość i prostotę. Jednak schematy te są często sprzeczne z danymi filogenetycznymi. Ponadto hierarchie i rangi tradycyjnej systematyki Linneusza (tj. Gromada, klasa, porządek, rodzina, rodzaj, gatunek) nie zawsze dobrze pasują do mikroorganizmów i często trudno jest zdecydować, który poziom hierarchii jest najbardziej odpowiedni dla dowolnej grupy pierwotniaków.
Wcześniej wiele systemów taksonomicznych definiowało pięć królestw życia: prokarionty (bakterie), protisty, rośliny, grzyby i zwierzęta. W tych schematach pierwotniaki są częścią Protista wraz z jednokomórkowymi glonami, okrzemkami, lęgniowcami i śluzowcami. Jednak grupa protistów zawsze była niezadowolona. Wynika to po części z tego, że protista jest częściowo definiowany przez kryterium negatywne. Innymi słowy, organizmy, które nie pasują do pozostałych czterech królestw, są domyślnie protista. Ponadto niektóre protisty są filogenetycznie bliżej spokrewnione z pozostałymi trzema królestwami eukariotycznymi niż z innymi protistami, a zatem protisty są wyraźnie polifiletyczne.
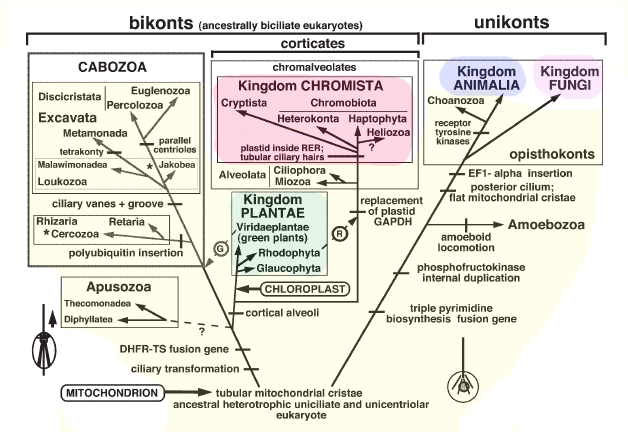
Cavalier-Smith proponuje pięć królestw eukariotycznych składających się z podstawowych, a więc parafiletycznych, królestw pierwotniaków i czterech królestw pochodnych: Animalia, Fungi, Plantae i Chromista (ryc.). Niektórzy z byłych protistów są teraz włączeni do królestw pochodnych, z którymi są najbliżej spokrewnieni, co powoduje, że królestwo Protozoa stało się monofiletyczne. Jednak pęcherzyki płucne, które obejmują wiele tradycyjnych pierwotniaków, takich jak apikomplexa, orzęski i bruzdnicowate, tworzą klad z chromistą, ale mimo to są umieszczane w królestwie pierwotniaków. Tak więc chromista nie jest holofiletyczny. Przeniesienie pęcherzyków płucnych do nowego królestwa zwanego Chromalveolata rozwiązałoby ten problem. Jednak prawdopodobnie spowodowałoby to kontrowersje i zamieszanie, ponieważ wiele pęcherzyków płucnych od dawna uważano za pierwotniaki.
Te strony są opracowywane i utrzymywane przez Mark F. Wiser z Tulane University (© 2000). Ostatnia aktualizacja 20 listopada 2018 r.